Molecular Detection of Complex Vertebral Malformation in Holstein Calves
By
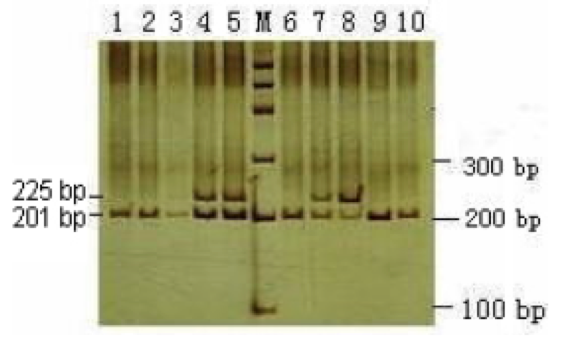
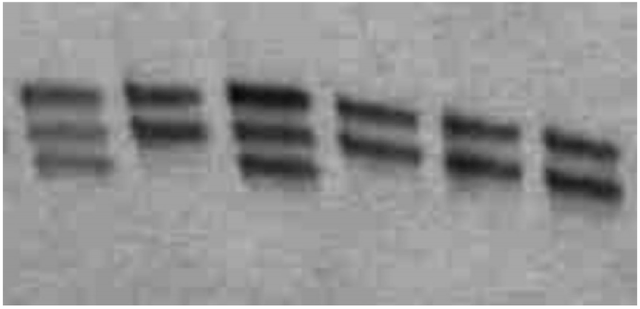
2014, Vol. 6 No. 05 | pg. 2/2 | « Current WorkThough carriers for CVM have been identified to be more desirable in the dairy market than are non-carriers in regards to milk production, recent research primarily focuses on carrier detection through molecular methods for the purpose of eventually eradicating the mutated SLC35A3 gene altogether, as well as studying the various attributes of the carriers. The ultimate purpose of these studies has been to identify quick and accurate detection methods to benefit the dairy market by reducing mortality in calves from CVM. Comparisons Between Carriers and Non-Carriers In order to better understand carriers of CVM and their potential importance in the economics of cattle breeding, Ghanem et al. (2008) compared the reproductive capabilities of CVM carrier Holstein cows in Japan with control groups. When analyzing the postpartum resumption of ovulation—the continuation of ovarian cycles after the birth of one calf—the researchers identified that there was no significant difference in the speed at which resumption occurs in carriers as compared to Holstein cows that were not carriers for CVM (Ghanem et al. 2008). However, CVM carrier cows were identified to have significantly lower conception rates than control cows, with the interval between calves observed at 463 days in carriers as compared to 399 days for control cows, suggesting the possibility of heightened mortality of embryos in CVM carrier cows (Ghanem et al. 2008). From an economic standpoint, this data is important because it provides invaluable data in regards to the reproductive capabilities of CVM carriers in comparison to non-carriers.However, due to the high mortality of calves with CVM, many researchers have focused on the importance of curbing the number of CVM carriers for the purpose of reducing the possibility of producing CVM calves. Recent estimates reveal that up to 30% of cows in several countries are heterozygous for CVM, a number that has encouraged those countries to attempt to reduce their frequency (Rusc et al. 2013). In order to reduce carrier frequency, research has focused on effective detection protocols for CVM carrier cows. By preventing the breeding of CVM carriers, the genetic frequency of the mutant SLC35A3 gene can be reduced and the dairy market can have consistently viable milk-producing offspring. Molecular Detection Methods of CVM Wang et al. (2011) provided one of the simplest techniques for the identification of CVM carriers, motivated by the fact that China’s dairy industry has become weakened by high abortion rates of calves due to genetic conditions such as CVM. Their Created Restriction Site PCR (CRS-PCR) protocol involved conducting PCR on the region of SLC35A3 that contains the potential transversion site using specific primers (Wang et al. 2011). These primers would produce a point mutation in the PCR products, ultimately resulting in the presence of a restriction site in non-carriers that the carriers for CVM lacked (Wang et al. 2011). After purification of the PCR products, Wang et al. (2011) used restriction fragment length polymorphism (RFLP). Due to the presence of the restriction site only in the non-carrier PCR product, the non-carriers produced two fragments and the carriers produced three on a gel in a restriction digest (Figure 3). To verify these results, the researchers used PCR data to identify the presence of two different alleles on the transversion site of the SLC35A3 gene versus just one in non-carriers. The presence of two alleles in some cows reveals that one of the two SLC35A3 genes underwent the transversion from guanine to thymine, consequently classifying those cows as carriers.
Figure 3: Created Restriction Site PCR (CRS-PCR) used restriction fragment length polymorphism (RFLP) results for Holstein bull SLC35A3. Lane M is a 100bp ladder marker. Carriers produced three bands in RFLP analysis at 225 bp, 201 bp, and 100 bp (the third band is not clearly visible) while non-carriers produced only two bands at 201 bp and 100 bp (From Wang et al. 2011). Wang et al. (2011) concluded that the CRS-PCR procedure for identifying CVM carriers and consequently removing those carriers from the breeding pool is the simplest and most cost-effective method among the available options. Despite this, there are several alternative methodologies that have since been developed to identify carriers. Mahdipour et al. (2010) used a PCR-Primer Induced Restriction Analysis (PCR-PIRA) in Karan Fries bulls to identify carriers for CVM. Using a different set of primers from Wang et al. (2011), the PCR-PIRA primers add a restriction site to the SLC35A3 gene DNA of unaffected bulls that is closer to the expected transversion site, allowing for a more focused approach to identify the single nucleotide conversion (Mahdipour et al. 2010). After PCR of SLC35A3 gene fragments, RFLP analysis revealed a single band for unaffected bulls and two distinct bands for CVM carriers (Figure 4). As was the case with the protocol of Wang et al. (2011), the identification procedure was verified through sequencing of the PCR products. Though the methodology was applied to a different type of cattle, a similar protocol can be applied to the SLC35A3 gene of Holstein cattle as well.
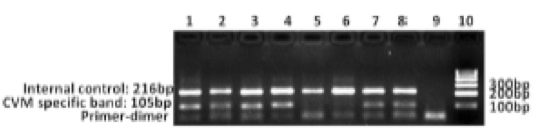
Figure 4: PCR-Primer Induced Restriction Analysis (PCR-PIRA) results in Karan Fries bull SLC35A3. Carriers produced two bands at 233bp and 212bp while non-carriers produced only one band at 233bp (From Mahdipour et al. 2010). PCR-PIRA as a means to identify CVM carriers in Holstein cattle has received both positive responses from later researchers as well as claims that the procedure is inaccurate and not ready to be applied to the dairy industry. Kotikalapudi et al. (2013) used the PCR-PIRA method on Holstein cattle to confirm the work of Mahdipour et al. (2010) on Karan Fries bulls. As was expected, the PCR-PIRA protocol as applied to Holstein cattle was successful in identifying CVM carriers and was confirmed with sequencing of PCR products. However, Kotikalapudi et al. (2013) also identified from the sequence analysis that one of the Holstein bulls that was a carrier for CVM also possessed additional mutations near the site of the transversion that results in CVM. The mutated area converted a thymine-guanine to a cytosine-thymine without any apparent phenotypic abnormalities in the bull. From this, it was deduced that the SLC35A3 gene is not as well understood as was previously believed, opening the door to more research on the gene and its abnormalities. While Kotikalapudi et al. (2013) confirmed the value of PCR-PIRA in identifying carriers for CVM, Wang et al. (2012) claimed that the procedure was not entirely advantageous because it still required the more complicated RFLP analysis in order to distinguish carriers from non-carriers. As a result, a new analysis, the Mismatch Amplification Mutation Assay PCR (MAMA PCR), was tested on several Holstein bulls (Wang et al. 2012). MAMA PCR has been an effective means of targeted amplification via primers with mismatches, a method that does not require RFLPs to identify point mutations such as transversions (Cha et al. 1992). Because MAMA PCR allows the amplification of one allele over another, Wang et al. (2012) were able to identify CVM carriers. Based on the PCR data alone, the study identified CVM carriers based on the presence of a 105bp band in addition to a control band (Wang et al. 2012). Non-carriers did not have the amplification of the specific allele of SLC35A3, and exhibited only the control band (Figure 5) (Wang et al. 2012). This study was particularly groundbreaking in CVM research because it allowed for the quick, cheap, and efficient molecular detection of carriers for the purpose of reducing the mutant gene frequency in the dairy market.
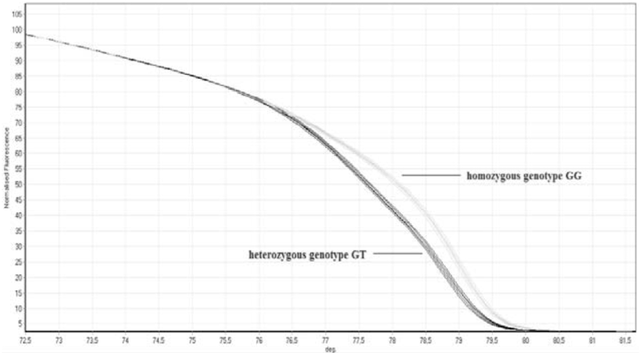
Figure 5: Mismatch Amplification Mutation Assay PCR (MAMA PCR) for Holstein bull SLC35A3. Carriers produced two bands (one control and one at 105bp), while non-carriers only produced the control band of 216 bp (From Wang et al. 2012). Gabor et al. (2012) developed a high resolution melting analysis (HRMA) of CVM that enabled them to conduct the entire detection experiment in a in a single step, without any need for further analysis of the PCR products, as was the case with previous PCR approaches. After conducting standard PCR on a portion of the SLC35A3 gene that contains the transversion characteristic of CVM for several Holstein calves, the genome samples were heated, cooled, and melted to produce a melting curve. The samples were already previously identified to be of either CVM carriers or non-carriers through PCR-PIRA, so Gabor et al. (2012) compared the melting curves of the two different sample types, finding consistent and significantly lower melting points for the carrier samples than non-carrier samples (Figure 6). The HRMA is a beneficial tool in the dairy market because it allows for a relatively quick and easy detection of CVM carriers.
Figure 6: High resolution melting analysis (HRMA) of SLC35Ain Holstein calves. Carriers exhibited a lower melting point for PCR products than non-carriers (78.15°C vs 79.42°C, respectively) (From Gabor et al. 2012). Application to the Dairy Market Most recently, Rusc et al. (2013) developed a program to decrease carriers for CVM, effectively using a relatively recent PCR-based approach known as PCR Single Stranded Conformation Polymorphism (PCR-SSCP). The methodology was first tested on CVM carriers and relies on recognizing the difference between carriers and non-carriers. After conducing PCR on the portion of the SLC35A3 gene that contains the mutation site for CVM, the researchers performed electrophoresis on polyacrylamide gel (Rusc and Kaminski 2007). Verified through more established PCR methods, Rusc and Kaminski (2007) identified consistent differences in electrophoresis results, with all carriers revealing three bands and non-carriers revealing only two bands on the gels (Figure 7).
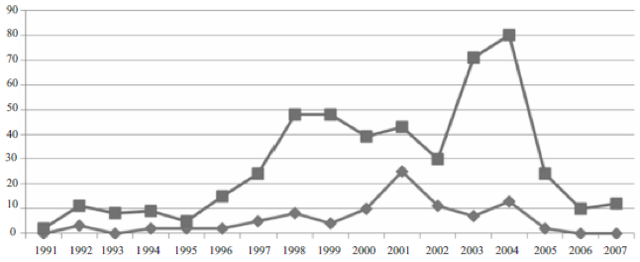
Figure 7: PCR Single Stranded Conformation Polymorphism (PCR-SSCP) for Holstein bulls. The first and third lanes represent carriers, while all other lanes represent non-carriers. Carriers produced three bands and non-carriers produced only two bands on the gel. Specific bp values were not provided (From Rusc and Kaminski 2007). With the success of PCR-SSCP in detecting CVM carriers, Rusc et al. (2013) applied their findings to the dairy market to reduce the frequency of CVM carriers in Poland. Using PCR-SSCP on 1,823 Holstein-Friesian bulls, the researchers found that nearly 17% of bulls were CVM carriers, leading them to screen all bulls that breeders brought into insemination sites for CVM to prevent carriers from reproducing and passing on the allele (Rusc et al. 2013). The direct intervention produced an eradication of CVM carriers born in 2006 and in subsequent years, suggesting that the detection of CVM carriers is both invaluable to the dairy market and fast-acting in reducing the CVM allele frequency (Figure 8). Rather than focusing on new methods of CVM detection, Rusc et al. (2013) stressed the importance of actually intervening in the dairy market to make important reductions in CVM carrier cattle.
Figure 8: Change in CVM carrier frequency in bulls, by birth year. The line with diamonds represents bulls who are carriers for CVM, while the line with squares represents non-carriers for CVM (From Rusc et al. 2013). Future DirectionsRegardless of the methodology used, it is clear that detecting carriers for CVM in Holstein cattle is critical to ensuring that no more calves are born with CVM or aborted as a result of the syndrome. Due to the high prevalence of CVM carriers in cattle populations globally, it is critical to focus research on two primary aspects: Perfecting the molecular approaches to CVM carrier detection and using those approaches to check and reduce the frequency of those carriers within dairy markets globally. Gabor et al. (2012) claimed that HRMA is effective in detecting CVM carriers under most circumstances, but the analysis is sensitive and has caused false positives. If HRMA, a highly efficient molecular approach to identifying the transversion characteristic of the mutated SLC35A3 gene, is to be applied successfully to the dairy market, it must be further perfected to ensure that false positives are not obtained and that a standardized procedure is established (Gabor et al. 2012). Furthermore, once those molecular approaches to CVM carrier detection are absolute in their efficiency, research should focus on the widespread use of those approaches in the dairy markets themselves. Because Hostein cattle are interbred at high rates, a wide-scale detection of CVM carrier frequencies will enable the establishment of longstanding programs to continuously remove carriers from the breeding population (Rusc et al. 2013). In addition to perfecting the molecular approaches for CVM detection and assessing the frequency of the mutated gene in cattle populations, future research should also focus on the effect of other mutations on the SLC35A3 gene. The presence of other mutations near the site of transversion within the SLC35A3 gene, without any overt phenotypic abnormalities, suggests that this gene should be further explored to better ascertain the various mutations that are possible and assess any less apparent effects on calves. Finally, research on CVM in Holstein calves should continue to assess the various attributes that CVM carriers and why those attributes exist. Holstein cows that are carriers for CVM are better milk producers but have lower conception rates than non-carriers, suggesting that there are a variety of resulting effects the single transversion mutation has on cattle (Qin et al. 2010; Ghanam et al. 2008). Understanding the molecular genetics and biochemical pathways that the mutation has on carriers may allow for therapies to improve the dairy market by producing highly viable offspring. ReferencesAgerholm, J. S., Bendixen, C., Ambjerg, J., & Anderson, O. (2004). Morphological variation of “complex vertebral malformation” in Holstein calves. Journal of Veterinary Diagnostic Investigation, 16, 548-553. Agerholm, J. S., Bendixen, C., Andersen, O., & Ambjerg, J. (2001). Complex vertebral malformation in Holstein calves. Journal of Veterinary Diagnostic Investigation, 13, 283-289. Cha, R. S., Zarbi, H., Keohavong, P., & Thilly, W.G. (1992). Mismatch Amplification Mutation Assay (MAMA): Application to the c-H-ras Gene. Genome Research, 2, 14-20. Gabor, M., Miluchova, M., Anna, T., Zuzana, R., & C. J. (2012). Detection of complex vertebral malformation carriers in Slovak Holstein cattle by high resolution melting analysis. Acta Veterinaria, 62, 239-248. Ghanem, M. E., Isobe, N., Kubota, H., Suzuku, T., Kasuga, A., & Nishibori, M. (2008). Ovarian Cyclicity and Reproductive Performance of Holstein Cows Carrying the Mutation of Complex Vertebral Malformation in Japan. Reproduction of Domestic Animals, 43, 346-350. Kotikalapudi, R., Patel, R.K., Sunkara, P.S.S., & A. Roy. (2013). Detection of Silent Homozygous Polymorphism in Exon 4 of SLC35A3 Gene in a Holstein Cattle Carrier for Complex Vertebral Malformation. International Journal of Veterinary Science, 2, 61-64. Mahdipour, M., Sharma, A., Dubey, P.P., Kumar, V., Misra, B., & Singh, A. (2010). Identification of Complex Vertebral Malformation using Polymerase Chain Reaction–Primer Introduced Restriction Analysis in Karan Fries Bulls. Current Trends in Biotechnology and Pharmacy, 4, 850-854. Nielsen, U. S., Aamand, G.P., Andersen, O., Bendixen, C., & Nielson, V.H. (2002). Effects of complex vertebral malformation on fertility traits in Holstein cattle. Livestock Production Science, 79, 233-238. Patel, R. K. (2012). Complex vertebral malformation (CVM) in Holstein. Wayamba Journal of Animal Science, 4, 483-485. Qin, C., Zhang, Y., Dong-Xiao, S., Yung, Y., & Ya-Chun, W. (2010). Identification of the complex vertebral malformation gene in Chinese Holstein and its association with dairy performance traits. Hereditas, 32, 732-736. Rusc, A., & Kaminski, S. (2007). Prevalence of complex vertebral malformation carriers among Polish Hosltein-Friesian bulls. Journal of Applied Genetics, 48, 247-252. Rusc, A., Hering, D., Puckowska, P., Barcewicz, M., & KamiÅ„ski, S. (2013) Screening of Polish Holstein-Friesian bulls towards eradication of Complex Vertebral Malformation (CVM) carriers. Polish Journal of Veterinary Sciences, 16, 579-581. Thomsen, B., Horn, P., Panitz, F., Bendixen, E., & Peterson, A. H. (2006). A missense mutation in the bovine SLC35A3 gene, encoding a UDP-N-acetylglucosamine transporter, causes complex vertebral malformation. Genome Research, 16,: 97-105. Wang, C., Tong, Q., Hu, X.Z., Yang, L. G., & Xhong, X. Q. (2011) Identification of complex vertebral malformation carriers in Holstein cattle in south China. Genetics and Molecular Research, 10,: 2443-2448. Wang, S., Hao, H., Zhao, X., Zhu, H., & Du, W. (2012). A rapid mismatch polymerase chain reaction assay to detect carriers of complex vertebral malformation in Holstein cattle. Journal of Veterinary Diagnostic Investigation, 24, 568-571. 
From the Inquiries Journal Blog ")  Related ReadingMonthly Newsletter SignupThe newsletter highlights recent selections from the journal and useful tips from our blog. Suggested Reading from Inquiries Journal
Inquiries Journal provides undergraduate and graduate students around the world a platform for the wide dissemination of academic work over a range of core disciplines. Representing the work of students from hundreds of institutions around the globe, Inquiries Journal's large database of academic articles is completely free. Learn more | Blog | Submit Follow IJ
Latest in Biology |