A Study of Planarian Phototaxis, Tactile Response and Regeneration
By
2014, Vol. 6 No. 03 | pg. 1/2 | »
IN THIS ARTICLE
KEYWORDS
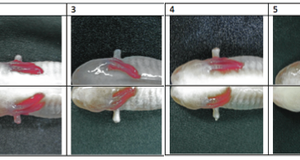
IntroductionThe phylum Platyhelminthes is comprised of aquatic worms that span over 20,000 species of either free-living flatworms or parasitic flukes and tapeworms (Rupert et al, 2004). The class Turbellaria consists of the free-living flatworms that are the focus of this study. Specifically, the negative phototaxis behavior, regeneration phenomenon, and tactile response of the Dugesia flatworm were experimentally assessed. The Dugesia flatworm (also known as a planarian), along with all worms in the Platyhelminthes phylum, is an acoelmate with no blood vascular system, making the worm an anatomically simple organism that relies on diffusion for all its transport needs. The most notable anatomical features of the planarian are a ciliated epidermis, a centralized brain with nerve chords extending throughout the rest of the body, various muscle fibers responsible for movement, and a ventrally-located mouth with a protrusible pharynx that brings algae, bacteria, and protozoans into a digestive syncytium (Rupert et al, 2004). Though they possess a highly simplified anatomy, planarians still exhibit certain complex behaviors. For one, planarians reveal a negative phototaxis behavioral response, meaning that they evade light (Inoue et al, 2004), and this response may be the result of a variety of factors. The two eyes in planarians are made up of only pigment cells and photoreceptor cells, and it is these photoreceptor cells that respond to light, although there are a number genes that seem to be responsible for this negative phototaxis. One gene, DjGAD, seems to be a likely candidate gene; DjGAD-positive planarians exhibit negative phototaxis while DjGAD-knockout planarians do not. DjGAD (the enzyme it produces is GAD, or glutamic acid decarboxylase) seems to be associated with GABA neurotransmitter production, so the presence of the enzyme and its product GABA seem to be associated with the phototaxis function in planarians (Umesono and Agata, 2009). RNA interference (RNAi) studying have also been used to identify the Djsnap-25 gene (which codes for the SNAP-25 protein) as another likely candidate, showing that planarians that were unable to express Djsnap-25 had impaired phototaxis behavior (Takano et al., 2007). Furthermore, immunostaining studies have also confirmed that negative phototaxis is temporarily impaired after decapitation of planarians, suggesting that regeneration of the photoreceptor system occurs during head regeneration (Inoue et al, 2004).Secondly, regeneration, the re-formation of decapitated parts, in planarians has been the focus of many studies of similar phenomena in other organisms. In planarians, regeneration occurs through the division and differentiation of pluripotent stem cells—neoblasts—that migrate to the regenerative bud after decapitation (Newmark, 2000). Like with the negative phototaxis response, regeneration behavior in planarians seems to be regulated by a number of factors. Using RNA interference studies, former researchers have found that β-Catenin (a protein that can control transcription and cell adhesion) acts as a sort of stimulator of proper planarian regeneration; in the absence of β-Catenin, decapitated planarians formed heads at their posterior ends as opposed to tails (Gurley et al, 2008). Other researchers identified the candidate gene smed-2 as critical to the regeneration process, without which regeneration did not occur in planarians (Reddien et al, 2005). With regards to the tactile response in planarians, there is sparse mechanistic or molecular information in the literature regarding the phenomenon. Generally speaking, planarians exhibit tactile sensitivity, and this sensitivity has been attributed to the anteromedian portion of the head (consisting of mucus, microvilli, and gland cells that form a complex with muscle cells and nerve endings) (Farnesi and Tei, 1985). It has also been found that the various sensory organs that are linked to the central brain via sensory nerves are responsible for the tactile responses exhibited in planarians (Cebria, 2007). This paper describes the experimental observations of phototaxis, tactile response, and regeneration after decapitation for a Dugesia flatworm. Based on prior literature, it was hypothesized that the planarian would exhibit negative phototaxis (movement away from source of light), tactile sensitivity resulting in movement away from source of touch, and a complete regeneration into two planarians approximately 5 to 6 days after decapitation. MethodsA live Dugesia flatworm was obtained from a beaker of stock culture and added to a petri dish with pond water. The stretched length of the flatworm in millimeters was obtained so as to assess the change in length from pre-decapitation to post-decapitation. Using a pipette tip, the planarian was gently touched on both posterior and anterior ends to assess response behavior. Finally, the petri dish was placed under a light microscope to assess the phototaxis response of the planarian. After the measurements for length and observations of phototaxis and tactile responses, the planarian was placed in ice for approximately 5 minutes to slow its movement and then decapitated slightly anterior to the mouth. The head and tail regions were placed in separate, labeled petri dishes to confirm, after regeneration, from which region the regenerated planarian was derived. Measurements of the head and tail region were taken immediately taken after decapitation, 30 minutes after decapitation, and 7 days after decapitation. Between measurements, the labeled petri dishes were incubated at room temperature, and the water in the petri dish was changed every three 3 to 4 days. After the final measurements, each of the regenerated planarians was finally returned to the stock culture. ResultsThe “parent” planarian obtained for this experiment from the stock culture was observed for phototactic and tactile response behaviors. When placed under a light microscope, the planarian attempted to move away from the light source. This was repeated several times to ensure that the planarian exhibited the same response each time. Because the planarian moved away from the light source, it exhibited negative phototaxis. Additionally, when touched on both its posterior and anterior ends with a pipette tip, the planarian attempted to move away from the tip, exhibiting sensitivity to touch. Table 1
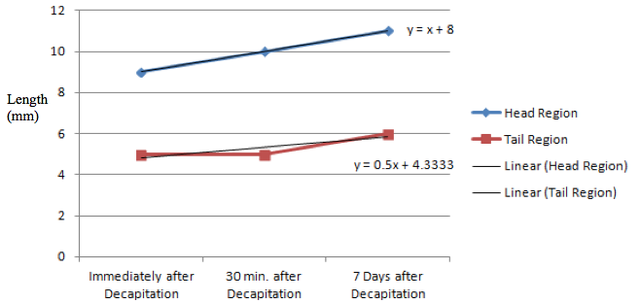
With regards to regeneration itself, the planarian length before decapitation was 14 millimeters and each individual region grew to 11 millimeters (head region) and 6 millimeters (tail region) after 7 days of incubation at room temperature in pond water. The average rate of growth over 7 days, as extrapolated from linear regression lines, of the head region was double that of the average rate of growth of the tail region. Figure 1: Length of the planarian at various points after decapitation. Note that the original length of the planarian was 14 millimeters. 
From the Inquiries Journal Blog ")  Related ReadingMonthly Newsletter SignupThe newsletter highlights recent selections from the journal and useful tips from our blog. Suggested Reading from Inquiries Journal
Inquiries Journal provides undergraduate and graduate students around the world a platform for the wide dissemination of academic work over a range of core disciplines. Representing the work of students from hundreds of institutions around the globe, Inquiries Journal's large database of academic articles is completely free. Learn more | Blog | Submit Follow IJ
Latest in Biology |