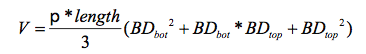From Discussions VOL. 5 NO. 2Effects of Simulated Deer Browsing Intensity and Fragmentation on Regenerative Dynamics of Acer saccharum Marsh. (Sugar Maple) Plants in Temperate Forests
IN THIS ARTICLE
KEYWORDS
AbstractThe deer browsing effects on the regeneration of woody plants in forested ecosystems is not fully understood. More can be learned by clipping plants in specific ways to mimic some aspect of a deer browsing regimen, especially intensity. Additionally, browsing effects are not understood in the context of fragmented forests. A fragmented forest has a "forest interior" and a "forest edge," which is the area closer to the forest edge. In this study, we simulated low and high levels of natural deer browsing intensity via "clipping" of Acer saccharum Marsh. (sugar maple) in three fragmented forests throughout northeastern Ohio, USA. At each site, 15 plants were randomly selected in forested habitats within 40 m (forest edge) as well as 40-100 m (forest interior) from the forest edge. In each habitat, leaves of plants were either (1) clipped at high intensity (0.9 g of biomass), (2) low intensity (0.3 g), or (3) no intensity (i.e. no clipping). Plant height, aerial crown cover, and basal diameter as well as numerous branch characteristics (length, diameter, leaf and bud production) were measured before and over two years following clipping treatment. The results showed that the simulated deer browsing intensities did not influence the response variables. They also showed that leaf and bud production as well as total volume of plants were greater in forest edge habitats than in forest interiors. Our results suggest that natural levels of deer browsing intensity do not interfere with growth dynamics of juvenile sugar maple plants and that the forest edge has a higher rate of growth than forest interiors despite the old age (~ 100 years) of forest edges. IntroductionIn forested ecosystems throughout the eastern United States, white-tailed deer (Odocoileus virginianus Zimmerman) is a common predator of understory woody plant species (Johnson et al. 1995). Over the last 100 years, some of these forests have experienced dramatic increases in deer densities as a result of altered hunting regulations and anthropogenic removal of deer predators (Russell et al. 2001). Increasing deer densities have led to concerns about over-browsing of woody plant species (Russell et al. 2001). Excessive browsing of juvenile woody plants, in particular, can prevent young canopy trees from transitioning from smaller to larger size classes to complete their natural life histories (Harmer 2001). The deer browsing effects on the regeneration of woody plants in forested ecosystems is not fully understood. More can be learned by clipping plants in specific ways to mimic some aspect of a deer browsing regimen, especially intensity. (Danell et al. 1994). However, most simulated deer clipping experiments are not realistic because deer browsing regimens were not accurately characterized. One such characteristic of the browsing regimen commonly misunderstood is browsing intensity. Other studies have used low levels of simulated browsing intensity (25-30% removal of green biomass via clipping) to increase plant growth (Bergstrom and Danell 1987) and higher levels (75-100%), in contrast, to slow growth and lower survivorship (Puetmann and Saunders 2001). However, this use of percentages rarely reflects natural levels of deer browsing intensity. Also, these studies are not conducted in natural settings, so interactive effects involving browsing and other environmental factors are not implemented. One such environmental factor is fragmentation. Forest fragmentation, due to anthropogenic disturbance, has increased the total area of forest habitats with forest edges over 100 years of age (Chen et al. 1992, Saunders et al. 1991). Compared to forest interiors, forest edges can have different levels of resources (e.g., light) (Euskirchen et al. 2000) altering plant growth and regeneration (Matlack 1994, Murcia 1995). How interactive effects like deer browsing intensity and forest fragmentation influence the regenerative dynamics of young woody plants has been poorly understood. Our objective was to evaluate the effects of deer browsing intensity on juvenile plant growth, using Acer saccharum Marsh. (sugar maple) in fragmented forests of northeastern Ohio. We administered three different intensities of deer browsing via clipping, and evaluated plant growth patterns over 3 years. We sought to answer two questions: (1) Do juvenile sugar maples respond differently to deer browsing at various intensities? (2) If so, do these responses differ between juvenile sugar maples located in forest edge habitats versus forest interiors? METHODSStudy SitesOur study was conducted at three different sites near Cleveland, Ohio. Both Bole and Firman Woods are located at The Holden Arboretum, Geauga and Lake Counties (41°36'N, 81°18' W; 8645 ha; 244 m above sea level), and the third site is located at Furnace Run Metro Park (41°15' N, 81°36' W; 360 ha; 340 m above sea level), which is part of the Summit County Metro Parks System. Regional temperature averages 10.8 °C, and mean monthly maximum and minimum temperatures occur in July (23.4 °C) and January (-2.7 °C), respectively (1971-2000; NCDC 2004). Annual rainfall is 91.6 cm on average, of which 60% occurs during the growing season from April through September. The three forested sites are situated on the Glaciated Allegheny Plateau of the Beech-Maple Forest Region (Braun 1950). They are second-growth temperate forests with a canopy composed of mainly sugar maple along with Fagus grandifolia Ehrh., Fraxinus americana L., Carya Spp., Prunus serotina Ehrh., and Acer rubrum L. In the understory, sugar maple, Fraxinus americana L., and Lindera benzoin Blume. predominate. Our sites have straight, north-facing edges that are ≥ 100 m long. Furnace Run (Summit County, 41°15' N, 81°37' W; 156 ha) is located along a residential road, Bole Woods (Geauga County; 40° 57' N, 82° 28' W; 200 ha) is situated next to an abandoned agricultural field, and Firman Woods (Lake County; 41° 37' N, 82° 18' W; 170 ha) is adjacent to a privately owned and maintained field. Aerial photographs suggest these edges are ~100 years old. All three forests are on gentle (2-6%) to moderately steep (6-15%) slopes and have deep soils (depth of 105-150 cm) and acidic subsoils (Williams and McCleary 1982, Ritchie and Reeder 1991, USDA 2008). The poorly drained soils that characterize Bole Woods are mostly Platea silt loams on medium textured glacial till with a silt loam surface and a silty clay loam substratum (Williams and McCleary 1982). They also contain a fragipan layer where the water table above is seasonally perched. The moderately drained soils at Firman Woods have a seasonally high water table, but are Ellsworth silt loams with a shale substratum (Ritchie and Reeder 1991). In contrast, the soils in Furnace Run overlay a silty clay loam substratum and are characterized as a Mahoning silt loam formed from glacial till (USDA 2008). Data CollectionData were collected over three years (2006-2008) from individually tagged sugar maple plants. In June and July of 2006, 30 juvenile plants between 30 cm and 1 m tall were randomly selected at each of the three forest habitat sites. 15 plants were selected < 40 m from the forest edge, and 15 were selected in the forest interior (40-100 m from an edge). Following plant selection, pre-clipping data were collected. Each plant was measured for height and basal diameter of the main stem. We then measured each branch off the main stem for length, diameter (at each end of a branch), as well as the number of buds and leaves. Additionally, each branch was marked with a uniquely colored piece of phone wire to keep track of the changes in these characteristics for each branch over time. A branch was defined as any living woody stem > 2 cm in length or any woody stem < 2 cm but with a living woody sub-branch > 2 cm long. Branch diameter was measured using digital calipers (accurate to ±0.01 mm; Mitutoyo Series 500, Kanagawa, Japan). Browsing intensity via clipping was administered in July 2006. In each forest habitat, terminal branches from each plant were randomly selected and received either one of three intensity clippings: (1) low-intensity involving the removal of 2-4 leaves (~0.3 g dry biomass) from a branch, (2) high-intensity or the removal of ~0.3 g of dry biomass from each of three branches (~0.9 g in total), or (3) no clipping. These clipping intensities differ from most simulated browsing studies because we did not defoliate our plants based on percentages. Instead, we simulated natural levels of deer browsing intensity. In other simulated browsing studies, researchers would remove a percentage of green biomass, ranging from 25 to 50% for low intensity (Cooper et al. 2003, Saunders and Puettmann 1999b) and 75 to 100% for high intensity (Canham et al. 1994, Saunders and Puettmann 1999a). These percentages work well for understanding plant responses to defoliation and heavy browsing. However, deers do not prefer sugar maple (Strole and Anderson 1992) and generally would not completely defoliate them. Thus, we clipped at intensities based on a typical bite size of deer (.3 g per bite) and the number of bites expected per deer browsing event (1-3 bites on average) (Shipley and Spalinger 1995). Following clipping, each plant was fenced to prevent natural deer browsing over the course of the study. Cylindrical-shaped fences were made of 2.54 cm poultry cloth and were 2 m tall and 1 m in diameter. Plants were re-measured in July 2007 and 2008. Statistical AnalysisData were analyzed in the context of a randomized block, multi-factorial experimental design with repeated measures and sampling, with simulated deer browsing intensity, location with respect to a forest edge, and time as fixed factors. At the plant level, we evaluated the effect of clipping on the collective response of unclipped branches of each plant. Response variables included total plant volume (calculated by summing all branch volumes), height to basal diameter ratio, and total number of leaves, buds, and branches per plant. Branch volume was calculated as the volume of a frustum using the following equation,
These data were analyzed using analysis of covariance (ANCOVA) with pre-treatment data used as the covariate. At the branch level, we examined mean branch volume and mean branch length involving all branches. These data was analyzed using ANOVA. Also, we compared those branches which were clipped in 2006 to randomly selected terminal branches from unclipped control plants and examined changes in the number of leaves and buds per branch. Branches that died in 2007 and 2008 were excluded from these analyses, and data were analyzed using ANCOVA. Analyses were performed using PROC MIXED in SAS 9.1 for Windows (SAS, 2001). When necessary, data were natural-log transformed to meet model assumptions; for all analyses, α = 0.05.Continued on Next Page » From the Inquiries Journal Blog   Related ReadingJournalQuest is a free program to help academic student publications increase online readership and distribution. If you are interested in enrolling a journal at your school, please visit the JournalQuest website. Monthly Newsletter SignupThe newsletter highlights recent selections from the journal and useful tips from our blog. Suggested Reading from Inquiries Journal
Inquiries Journal provides undergraduate and graduate students around the world a platform for the wide dissemination of academic work over a range of core disciplines. Representing the work of students from hundreds of institutions around the globe, Inquiries Journal's large database of academic articles is completely free. Learn more | Blog | Submit Follow IJ
Latest in Biology |