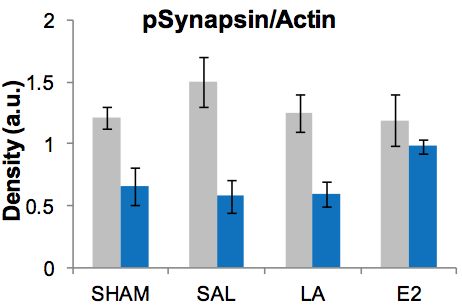
From Discussions VOL. 8 NO. 2The Contribution of HPG Axis-Hormones and Hormone Replacement Timing on Cognition and Associated SignalingResultsOverall, our data reveals trends in protein expression as the term of the treatment increases. Phosphosynapsin expression decreases with age regardless of the treatment. Phospho-CAMKII expression as opposed to total CAMKII expression varies with regards to treatment, while synaptophysin expression increases with age in each treatment group. Phospho-synapsin is not affected by ovariectomyor by treatments in the short term after ovariectomy, but it is reduced in the long term with each treatment. Interestingly, the reduction in phospho-synapsin expression from E2 replacement had a smaller difference compared to the SHAM mice and the other treatments (Figure 1). The trend of decreased phospho-CAMKII expression with increasing age is not as defined as with the other proteins because with the SAL treatment there is an overexpression of the protein long after the onset of menopausein particular 6 months after; however the autophosphorylation of CAMKII is not affected within 3 months from the OVX. LA increased levels of pCAMKII significantly at both time-points yet, levels in LA treated animals were significantly lower to those of OVX animals in the long-term group (Figure 2). The trend in synaptophysin expression is age-related upregulation independent of the groups being treated. The protein expression is not altered by OVX or any of the treatments. When LA treatment is started 4 months post-OVX, the downregulation of LH significantly increased synaptophysin expression beyond age-associated levels. However, the E2 treatment started 4 months after postOVX shows a smaller increase in protein expression when compared to the other treatments (Figure 3). Figure 1. Protein expression of phosphorylated synapsin. Short-term treatment shown in grey and long in blue. There is no effect of ovariectomy or any of the treatments when started short term after ovariectomy. E2 replacement started a long interval after ovariectomy significantly upregulates the levels of pSynapsin compared to all other groups.
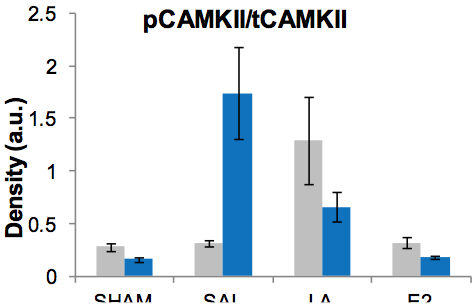
Figure 2. Protein expression of phosphorylated CAMKII. Short-term treatment shown in grey and long in blue. There is no effect of ovariectomy on pCAMKII expression; however, LA treatment leads to a significant upregulation compared to all treatments. pCAMKII is significantly upregulated by OVX when measured 6 months after the procedure compared to all groups. LA treatment when started long-term is significantly downregulated compared to short-term and long-term OVX but still significantly elevated compared to SHAM and E2 groups.
Figure 3. Protein expression of synaptophysin. Short-term treatment shown in grey and long in blue. There is no effect of ovariectomy or any of the treatments when administered shortterm after OVX. Synaptophysin expression is upregulated by age in all conditions (short vs. long). LA treatment when administered longterm after OVX leads to a significant increase in synaptophysin expression compared to all other groups.
Conclusion and DiscussionBased on the results, the hypothesis that cognitive changes after menopause are primarily driven by increases in LH, not decreases in estrogen levels, is partially proven because the effects of LA and E2 are very similar in protein expression immediately post-OVX, but different long after OVX. This tells us that E2 is not as effective as LA when applied in the long-term, hence proving that cognitive changes post menopause are driven by increases in LH. In addition, the data discloses that the tested signaling molecules are not considerably modified during menopause at least within the first 3 months. This suggests that cognitive impairment observed by OVX is unlikely to be driven through these cascades and are affected by some other cascades. Our results also show that synaptic transmission associated proteins show age-related effects in all our groups. For example, synaptophysin decreases with age in the hippocampus and various cortical regions [7]. Previously published data has shown that aging can lead to the incorrect regulation of many proteins involved in neuronal signaling that may be associated with age-related diseases [13]. This makes sense because protein levels can be regulated by hormonal control, but with time, incorrect hormone levels can result in the dysfunction of proteins involved in synaptic transmission. Dysfunction in transmission can lead to a decrease in neurotransmitter release. In another set of results, age-related downregulation of phospho-synapsin is prevented by E2 replacement when treatment is started long after OVX. The significance of these findings is unclear given that E2 replacement is not effective at improving cognition at this later interval. On the other hand, LA increases synaptophysin expression in our long-term group. It is unlikely that synaptophysin is a major modulator of cognition after menopause based on our data; instead, it might modulate the efficiency of synaptic vesicle cycles. However, the additional increases produced by LA could take part in cognitive protection. This could be due to LA being more effective than E2 in downregulating LH in the long term. Lastly, while pCAMKII is not substantially changed by OVX at 3 months, it is significantly increased at 7 months post OVX. This could be in response to the decrease in levels of neurotransmitter release post-OVX in the long term due to synapsin. The increase in pCAMKII allows more AMPA receptors to reach the synapse to accept the few neurotransmitters released. On the other hand, LA treatment increases pCAMKII when treatment is started immediately after and long-term compared to SHAM, but the magnitude of increase is significantly lower than that of OVX (longterm). This is reasonable because LA downregulates LH creating effects similar to SHAM and E2. It is challenging to isolate these signaling cascades to guarantee that other factors are not interfering with the processes. For example, calcium plays a huge role in cell signaling and transduction. CAMKII autophosphorylation is specific to intracellular calcium release, which is known to both improve cognition and lead to cytotoxicity. Differences in the magnitude of pCAMK in this study may therefore reflect positive and noxious increases in Ca++ release. The next steps would possibly be to test more samples, test other proteins in the signaling cascade to confirm the process and reasons why the levels go as the results show, or to somehow isolate these signaling cascades and see their effects purely from the treatments administered. AcknowledgementsI would like to thank my advisor, Michael Benard, and my mentor, Gemma Casadesus, both of whom advised me in what direction to go with my research. I would also like to thank Jaewon Chang, Hyun Jin Kim, Yoelvis Garcia, and Denise Hatala in assisting me with my proj- ect. This work was carried out with funds from NIA R01 AG032325-01. References
From the Inquiries Journal Blog   Related Reading JournalQuest is a free program to help academic student publications increase online readership and distribution. If you are interested in enrolling a journal at your school, please visit the JournalQuest website. Monthly Newsletter SignupThe newsletter highlights recent selections from the journal and useful tips from our blog. Suggested Reading from Inquiries Journal
Inquiries Journal provides undergraduate and graduate students around the world a platform for the wide dissemination of academic work over a range of core disciplines. Representing the work of students from hundreds of institutions around the globe, Inquiries Journal's large database of academic articles is completely free. Learn more | Blog | Submit Follow IJ
Latest in Biology |