From Discussions VOL. 5 NO. 1Turning Behaviors in the Cockroach Blaberus discoidalis
By
Discussions 2008, Vol. 5 No. 1 | pg. 1/1
IN THIS ARTICLE
KEYWORDS
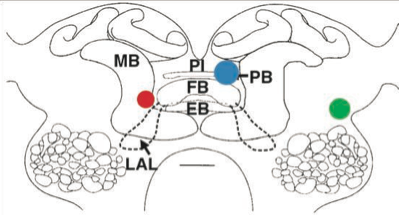
INTRODUCTIONLegged insects are able to traverse a wide range of unpredictable terrain and assess their surroundings by responding to sensory input absorbed through their eyes and antennae. When an antenna comes into contact with an obstacle, the cockroach changes gait patterns accordingly to navigate around the obstacle (Mu, 2005). In this study, discrete electrolytic lesions were used to assess the role of various regions in the brain in completing complex obstacle negotiation behaviors. One region in particular, the central complex, is of interest because it has been shown to have antennal inputs as well as effects on motor control (Ridgel et al 2007). Cockroaches were tested on a vertical and horizontal track in order to examine the differences in turning on a flat horizontal surface and turning on a vertical surface. Preliminary data suggests that lesions to the central complex result in turning deficits on both flat and vertical surfaces. MATERIALS AND METHODSAnimals Adult Blaberus discoidalis cockroaches were housed in small colonies in a twenty liter bucket. These were maintained in a 12-12 hour lightdark cycle at 25°C. Animals had ad libitum access to water and dry food in the form of chicken feed. Both male and female animals were used in these experiments. Horizontal climbing apparatusCockroaches walked through a U-shaped turn made of Plexiglas. The turn was placed on a glass plane with a mirror at a 45° angle below it to reflect images for video recording. Vertical climbing apparatusThe same Plexiglas U-shaped turn used in the horizontal climbing apparatus was mounted on a square wooden frame. The frame had a plastic screen stretched underneath the Pelxiglas structure, which the cockroaches used to climb. Preparation and surgeryIntact animals were taken and anesthetized with exposure to sub-zero temperatures for five to eight minutes. Their abdomens were then painted white with WhiteOut pen to aid in viewing their leg movements. Individuals were given at least one hour to recover. They were then run through the horizontal and vertical turns in order to determine each individual's normal behavior. After initial tests were completed, the cockroaches were anesthetized with cold again for five to eight minutes and placed into a stabilizing tube to prevent their heads from moving. Using a fine razor blade, the brain was exposed and a steel probe was inserted into the brain. A current of approximately 1.3 mA was applied for 30 seconds, and then wax was used to secure the removed exoskeleton to the head. All insects were given at least 24 hours to recover from surgery. HistologyFollowing the procedures of Ridgel et. al. 2007, the heads of each animal were removed after post-lesion tests were completed, then the brain was removed, and fixed in AAF (85% ethanol, 5% acetic acid, 10% formaldehyde). The brains were then dehydrated in graded ethanol series and embedded in Paraplast Plus (Fisher). Tissue blocks were sectioned at 12 μm and stained with 1% toludine blue. Sections were then viewed with a conventional light microscope. Data analysisAll videos were analyzed using WinAnalyze motion analysis software (Mikromak, Berlin). Videos were analyzed frame-by-frame and each trial was labeled either normal or abnormal based on pre-tests done with intact animals. Abnormal behaviors were defined as a permanent change in behavior from behaviors exhibited in the pre-lesion tests. RESULTSLesion locationsOf all the cockroaches tested, only four generated data suitable for analysis. The cockroaches had lesions on the top of the fan body, near the fan body, outside the central complex, and a sham as depicted in Figure 1. Figure 1. Drawing of the different locations of lesions. FB is the fan-shaped body, PB is the Protocerebral bridge, EB is the Ellipsoid body, MB is the Mushroom body, LAL is the Lateral accessory lobe, and PI is the Pars intercerebralis.
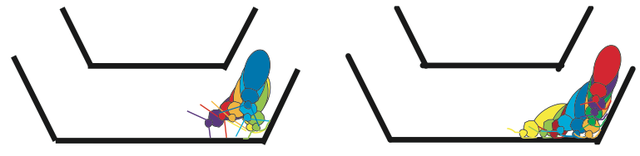
Turning behaviorsAbnormal turns were defined as a distinct change from an animal's pre-lesion behavior. Figure 2 is an example of an abnormal turn. The main difference was the lack of response to antennal feedback that resulted in the animal hitting the wall before turning. On both the horizontal and vertical turns, those cockroaches with lesions closer to the central complex were the source of a higher percentage of abnormal turns, see figure 3. The cockroach that had a lesion right above the fan-shaped body had an almost one hundred percent increase in abnormal turning behaviors. The lesion that was close to the central complex was approximately 52nm away, resulting in small increases in abnormal turning behavior. In the case of the lesion that was completely outside the central complex, the behavior patterns were similar to a sham. On the vertical turn, individuals often did not move which may be a result of a more difficult obstacle. Figure 2. Tracings of a cockroach going through a horizontal turn. The left picture shows a pre-lesion insect making antennal contact with a wall and responding to the sensory input. The right picture is a post-lesion insect that has no response to antennal contact with the wall and consequently only turns once its head comes into contact with the wall.
DISCUSSIONSensory input is extremely important in decision making for animals when they approach unfamiliar obstacles. By disrupting the flow of sensory information through the use of discrete lesions we can generate abnormal behaviors. The resulting behaviors and their severity are the result of lesion size and location of the lesion. Discrete lesions closer to the central complex resulted in a larger percentage of abnormal behaviors. The insect that had the lesion at the top of the central complex in the protocerebral bridge had an almost one hundred percent increase in abnormal behaviors. The protocerebral bridge is responsible for compensating in differences in left-right motor responses, and therefore regulating the swing-stance phases during turning (Strausfeld 1999). Sensory information from the legs offsets the effects of the lesion in the protocerebral bridge when the cockroaches came in contact with the wall. This feedback then caused the cockroach to turn, much like it did when it hit its head on the wall on the horizontal turn, lowering the level of abnormality on the vertical turn. The insect with a lesion completely outside of the central complex exhibited behaviors similar to a sham. Figure 3. Summary of the percent increase of abnormal behaviors depending on location of lesion and which obstacle the cockroach navigated. CC is the central complex.
In addition, the lesioned insects seemed reluctant to climb on the vertical turn. This could be the result of the inability to climb from lack of sensory input, or it may be a result of the lesioning process. However, once the insect managed to get on the screen, it was still able to climb. The overall conclusion made is that the closer a discrete lesion is to the central complex, the greater the increase in abnormal turning behaviors. It should be noted, again, that the insect is still able to navigate a vertical surface. Future StudiesBecause of the small sample size, this is merely a preliminary study. Conclusions about the effects of discrete lesions on the turns cannot be made with such little supporting evidence. By increasing the number of trials, we can make clearer conclusions about the effects of lesions on horizontal versus vertical turning. By localizing the lesions to a specific area, we can also increase the statistical significance of any findings made. AcknowledgementsI would like to thank Dr. Roy Ritzmann for his guidance throughout all my research. Cynthia Harley was also instrumental in teaching me the required techniques and helping me with my experiments. Al Pollack was a great help with the construction of our experiment. Last, Nicholas Courtney is a good friend and partner as we helped each other with our respective experiments. ReferencesMu L, Ritzmann RE (2005) Kinematics and motor activity during tethered walking and turning in the co croach, Blaberus discoidalis. J Comp Physiol A Neuroethol Sens Neural Behav Physiol 191:1037– 1054 Ridgel AL, Alexander BE, Ritzmann RE (2007) Descending control of turning behavior in the cockroach, Bla berus discoidalis. J Comp Physiol A 193:385–402 Ritzmann RE, Pollack AJ, Archinal J, Ridgel AL, Quinn RD (2005) Descending control of body attitude in the cockroach, Blaberus discoidalis and its role in incline climbing. J Comp Physiol A 191:253–264 Ritzmann RE, Ridgel AL, Pollack AJ (2008) Multi-unit recording of antennal mechano-sensitive units in the central complex of the cockroach, Blaberus discoidalis. J Comp Physiol A Strausfeld, N. J. (1999) A brain region that supervises walking. Prog Brain Res 123: 273‐84. From the Inquiries Journal Blog   Related ReadingJournalQuest is a free program to help academic student publications increase online readership and distribution. If you are interested in enrolling a journal at your school, please visit the JournalQuest website. Monthly Newsletter SignupThe newsletter highlights recent selections from the journal and useful tips from our blog. Suggested Reading from Inquiries Journal
Inquiries Journal provides undergraduate and graduate students around the world a platform for the wide dissemination of academic work over a range of core disciplines. Representing the work of students from hundreds of institutions around the globe, Inquiries Journal's large database of academic articles is completely free. Learn more | Blog | Submit Follow IJ
Latest in Biology |