From Earth Common Journal VOL. 5 NO. 1The Sixth Species Extinction Event by Humans
IN THIS ARTICLE
KEYWORDS
AbstractThe number of species becoming extinct has drawn a significant deal of attention from scientists and non-scientists alike. This research reviews recent literature citing evidence for the impact humans have had on our planet and how our biological systems are affected in both known species of flora and fauna as well as unknown species of flora and fauna, the latter lacking documentation as well as sightings by humans. Theoretical research is derived from previous research investigating the impacts of humankind's use of the land as well as population increases. Though there are many different definitions of what a mass extinction is and gradations of extinction intensity, a conservative approach is used to assess the seriousness of the current ongoing extinction crisis, setting the highest level of recognition for mass extinction, in extreme diversity loss associated with the Big Five extinction events (Barnosky, 2011). Understanding the relationship between extinction and functional diversity over time will be critical for making conservation work (Boyer & Jetz, 2014). If another mass extinction is allowed to progress, it would mean the end of biodiversity as we know it and would also mean that greater pressure would be placed on both humans and flora and fauna to survive in a world completely changed by the Anthropocene. Over the course of 8,000-10,000 years, humans grew in population and changed the landscape of the Earth (Foley, 2013). The research concludes that focus should be on preserving the environment and future research should be performed on the study of unknown species. IntroductionCurrently there is a great deal of attention being paid to the ever increasing effect humans have on the rate of extinctions on Earth. With our influence being a constant factor, species' extinctions are occurring very rapidly in biological systems globally. Recent discoveries suggest that a sixth mass extinction could be under way, given the number of known species losses over the millennia (Barnosky, 2011). Humankind's constant destruction of the environment has affected many different species of many ecosystems, but the majority can be categorized into two different groupings: Known Species and Unknown Species (Costello, 2013).
Over the course of 500 years, humans have caused extinctions of such speed and magnitude that it is rivaling the Big Five extinction events of the past. (Dirzo, 2014). Eventually a tipping point in time could be reached when Earth's changes will be rendered irreversible. Earth could reach an extreme within just a few centuries if current threats to many species are not alleviated (Pievani, 2014). There can be no doubt that the human race is the culprit behind these recent extinction events. The research came to conclude that the current period, known as the Palaeoanthropocene, is now a recognized period where humans are now an integral part of the Earth System rather than an external forcing factor (Foley, 2013). The purpose of this paper is to review the literature for evidence of how humankind is ultimately dooming many species of flora and fauna to extinction and how we are affecting our biological systems in terms of the number of known species that are going extinct as well as the unknown species that are going extinct. ResultsMass extinctions play a pivotal role in macroevolution. As a result of their frequency, speed, and global impact, mass extinctions have shaped global biodiversity throughout the history of life on Earth. Now as a result of human influence, both direct and indirect in nature, a serious extinction crisis could be possible in a matter of centuries. These activities include fragmentation of habitats, overpopulation, chemical pollution, invasive species, over-exploitation of resources in hunting, and overfishing. Species distribution is analyzed through variation of flora and fauna as well as habitable areas which would house such creatures. Defaunation patterns have also been investigated for evidence of future impacts of the Anthropocene. To prevent the impending sixth species extinction event, it is imperative that humans find ways to reduce the impact of such disasters not only for the future of every ecosystem on the planet, but also for the future of their descendants. For it is the children of the future that will inherit either the problems of their predecessors or the salvation their predecessors grant them (Pievani, 2014). One of the ways that humans can combat species extinction is to predict the number of species going extinct due to habitat loss. To make these predictions, biologists employ a mixture of species-area relationship (SAR) and species-fragmented relationship (SFAR) (Rybicki & Hanski, 2013). The SAR gives a quantitative description of the increasing number of species in a community with increasing area of habitat. In conservation, SARs have been used to predict the number of extinctions when the area of habitat is reduced. Such predictions are most needed for landscapes rather than for individual habitat fragments, but SAR-based predictions of extinctions for landscapes with highly fragmented habitat are likely to be biased because SAR assumes contiguous habitat. In reality, habitat loss is typically accompanied by habitat fragmentation. To quantify the effect of fragmentation in addition to the effect of habitat loss on the number of species, Rybicki and Hanski (2013) extend the power-law SAR to the species-fragmented area relationship. This model unites the single-species metapopulation theory with the multispecies SAR for communities. A realistic simulation model and empirical data for forest-inhabiting subtropical birds demonstrate that SFAR gives a far superior prediction than SAR of the number of species in fragmented landscapes. The results demonstrate that for communities of species that are not well adapted to live in fragmented landscapes, the conventional SAR underestimates the number of extinctions for landscapes in which little habitat remains and it is highly fragmented (Rybicki & Hanski, 2013).
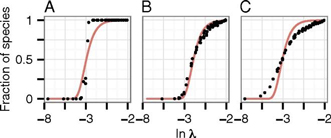
Fig. 1. Effect of Fragmentation on Species Number. As Figure 1 demonstrates, each of graphs A, B, and C show a different level of habitat fragmentation, with A showing the lowest effects of habitat fragmentation and C showing the highest effects of habitat fragmentation. The variation between each of the three cases is looked at. In graph A, there is no variation, while in B and C there is now tenfold variation in extinction and colonization rates, respectively. Graph A shows that with very high habitat cover, there are more species present as well as more individuals of each species present than in graphs B and C, which show lower habitat cover and thus a lower amount of species and individuals in each species present. The fragmentation effects on species number at landscape levels are due to local extinctions in habitat fragments and it is also because of nonviable metapopulations in highly fragmented landscapes. Fragmentation in very large spatial scales do not show the same conclusions because large habitat fragments hold individually viable populations and thus if the fragment sizes increase then extinction rates decrease (Rybicki & Hanski, 2013). The fraction of species persists in the simulation (points) and the value of P(λ) = exp (-b/λ) (continuous line) against the logarithm of metapopulation capacity. In (A), there is no variation among the species in any parameter (details in SI Text). In (B), parameter values were drawn from the same distributions as in (A), including roughly twofold variation in colonization and extinction rate parameters. In (C), the same parameter values as in (B) except that now there is tenfold variation in colonization and extinction rates. The fraction of species persisting in the simulation is the number of species persisting divided by 188 (200 in A), which is the maximum number of species surviving in landscapes with very high habitat cover (Rybicki & Hanski, 2013).
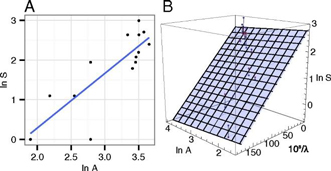
Fig. 2. Effect of Habitat Fragmentation on the Number of Tropical Bird Species. In Figure 2, each graph is a plot of the number of species of tropical birds against the number of available habitat area for each species. As the graph demonstrates, there is a positive correlation between the number of birds present and the level of forest cover; as forest cover decreases, so does the number of bird species that are present in these environments. Figure 2 illustrates the SAR and the SFAR in subtropical bird species in large (100-km2) forest landscapes with less than 40% native forest cover (n = 14). (A) The logarithm of species number against the logarithm of habitat area and (B) the logarithm of species number plotted against the logarithm of total habitat area and the inverse of the metapopulation capacity (1/λ). Note the orientation of the horizontal axes in (B), where the blue points give the actual values and the red points the projected values on the regression plane (Rybicki & Hanski, 2013). The habitats these unique tropical birds depend on are being depleted and slowly the birds are being driven to extinction because of humankind's growing presence (Rybicki & Hanski, 2013). Similar to other mass extinctions, the Anthropocene extinction event is affecting every taxonomic group in existence, even though some are clearly more effected by human influence than others. One conservative estimate implies that Earth could be losing approximately 11,000 to 58,000 species per year. (Mora, 2013) Many disappear before they can even be identified. Extinctions are also not as evenly distributed between taxonomic groups. For instance, amphibians are more affected by environmental changes than birds are. Many remaining species are also suffering from severe population declines as well. Terrestrial vertebrate populations have declined 25% and 67% of monitored invertebrate populations are currently declining by 45%. This loss of species from ecosystems, whether through local population declines or species extinction, will disrupt the natural function of ecosystems and humans will ultimately suffer from this impact because of the dependence on such vital ecosystems for survival and well-being (Dirzo, 2014). History shows that human interference has occurred since the end of the last ice age. This unique period is known as the Anthropocene, the era in which humans invented agriculture and became complex societies. Some would argue that this could be extended into well into the Pleistocene epoch (2.58 Ma) in a period known as the Palaeoanthropocene, an era that includes many of the first species in the genus Homo as well as the Holocene epoch and modern history. During the Paleoanthropocene, environmental and climate changes had a large impact on food supplies. Food supplies, in turn, were a determining factor in the migration and expansion of human populations. Agriculture production and the keeping of livestock gradually spread from community to community and this became important for emerging societies, whom depended on such resources for survival. New attitudes regarding to distribution of resources, stockpiling, territory, and work distribution also contributed to the first major population increase in human history. The increase in population density led to newer forms of interdependence between humans and nature and led to crop failure and floods. This often resulted in food shortages and, consequently, starvation. Yet with technological innovations, the human population increased, which increased the risk of subsistence crises (Foley, 2013). The Palaeoanthropocene involved gradual changes related to fire and land use. Furthermore, in this era, all species extinctions are related to regional effects. The Palaeoanthropocene has a diffused beginning that should not be distinguished by geological boundaries, as it is linked to local events and annual to seasonal timescales that are not recognized easily in terms of global assessments. The Anthropocene involved changes that stretched over the course of roughly 8,000 years. This resulted in more destruction of the natural world than at any other time in Earth's history. It has also marked the first time in Earth's history that a single species was wiping out more species because of a large increase in population numbers (Foley, 2013). As seen in Figure 3, during the Anthropocene there were significant developments in humankind that impacted Earth exponentially as technology improved and thus resulted in many species becoming extinct. In the Palaeoanthropocene, there is very minimal impact over the entire era due to the small number of individuals per species of hominids and the number of hominid species was far greater than it is currently (Foley, 2013).
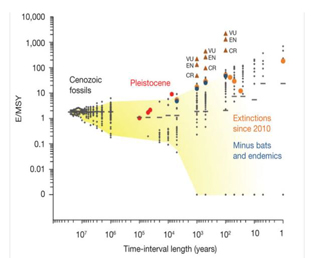
Fig. 3. Relationship between Extinction Rates and the Time Interval Over Which the Rates Were Calculated, for Mammals. Each small grey datum point in Fig. 3 represents the E/MSY (extinction per million species-years) calculated from taxon durations recorded in the Paleobiology Database (million-year-or-more time bins) or from lists of extant, recently extinct, and Pleistocene species compiled from literature (100,000-year-and-less time bins). More than 4,600 data points are plotted and clustered on top of each other. Yellow shading encompasses the ‘normal' (non-anthropogenic) range of variance in extinction rates that would be expected given different measurement intervals; for more than 100,000 years, it is the same as the 95% confidence interval, but the fading to the right indicates that the upper boundary of ‘normal' variance becomes uncertain at short time intervals. The short horizontal lines indicate the empirically determined mean E/MSY for each time bin. Large coloured dots represent the calculated extinction rates since 2010. Red represents the end-Pleistocene extinction event; Orange, the documented historical extinctions averaged (from right to left) over the last one, 30, 50, 70, 100, 500, 1,000 and 5,000 years; and blue the comparability of modern with fossil data by adjusting for extinctions of species with very low fossilization potential (such as those with very small geographic ranges and bats). For these calculations, ‘extinct' and ‘extinct in the wild' species that had geographic ranges less than 500 km2 as recorded by the International Union for the Conservation of Nature, all species restricted to islands of less than 105 km2, and bats were excluded from the counts (bats are under-represented as fossils, making up only about 2.5% of the fossil species count, as opposed to 20% of the modern species count). Brown triangles represent the projections of rates that would result if ‘threatened' mammals go extinct within 100, 500 or 1,000 years. The lowest triangle (of each vertical set) indicates the rate if only ‘critically endangered' species were to go extinct (CR), the middle triangle indicates the rate if ‘critically endangered' and ‘endangered' species were to go extinct (EN), and the highest triangle indicates the rate if ‘critically endangered,' ‘endangered,' and ‘vulnerable' species were to go extinct (VU). To produce Fig. 3, the last-occurrence records of Cenozoic mammals from the Paleobiology Database, as well as the last occurrences of Pleistocene and Holocene mammals from refs six, 32, 33 and 89-97 were determined. Researchers then used R-scripts (written by Nicholas Matzke) to compute total diversity, number of extinctions, proportional extinction, and E/MSY (and its mean) for time bins of varying duration. Cenozoic time bins ranged from 25 million to one million years. Pleistocene time bins ranged from 100,000 to 5,000 years, and Holocene time bins from 5,000 years to a year. For Cenozoic data, the mean E/MSY was computed using the average within-bin standing diversity, which was calculated by counting all taxa that cross each 100,000-year boundary within a million-year bin, then averaging those boundary-crossing counts to compute standing diversity for the entire million-year-and-over bin. For modern data, the mean was computed using the total standing diversity in each bin (extinct plus surviving taxa). This method may overestimate the fossil mean extinction rate and underestimate the modern means, so it is a conservative comparison in terms of assessing whether modern means are higher. The Cenozoic data are for North America, and the Pleistocene and Holocene data are for global extinction; adequate global Cenozoic data are unavailable. There is no apparent reason to suspect that the North American average would differ from the global average at the million-year timescale (Barnosky, 2011).Continued on Next Page » From the Inquiries Journal Blog   Related ReadingJournalQuest is a free program to help academic student publications increase online readership and distribution. If you are interested in enrolling a journal at your school, please visit the JournalQuest website. Monthly Newsletter SignupThe newsletter highlights recent selections from the journal and useful tips from our blog. Suggested Reading from Inquiries Journal
Inquiries Journal provides undergraduate and graduate students around the world a platform for the wide dissemination of academic work over a range of core disciplines. Representing the work of students from hundreds of institutions around the globe, Inquiries Journal's large database of academic articles is completely free. Learn more | Blog | Submit Follow IJ
Latest in Environmental Studies |